
跑了一段時間之後,你會開始聽到各種訓練主張。有人說 80% 的時間要跑 Zone 2,有人說挪威訓練法才是真的厲害,有人說 HIIT 才能最快進步。每個說法背後都有數據,每種訓練都有人跑出好成績。
問題不是哪個方法正確,而是這些訓練怎麼讓你變強的。
訓練是在給身體發訊號
我們在〈LT1、LT2 與三區模型〉裡談過 LT1 和 LT2,以及三區模型如何劃分訓練強度;〈心率區間三種測定方法〉則討論了如何以心率把這些區間邊界定下來。這篇往下一層——不是強度分區,而是每個強度區間在你身體裡觸發了什麼。
訓練的本質是刺激訊號。你施加一個夠強的壓力,身體偵測到這個壓力,啟動一連串的分子反應,在壓力消失之後進行適應。不同強度發出的訊號種類不同,觸發的適應也不同。Zone 2 跑步後和 HIIT 後,肌肉細胞裡激活的基因表現模式是不同的——這不是比喻,而是字面意思。
運動科學家 Andrew Coggan 在與 Hunter Allen 合著的《Training and Racing with a Power Meter》(VeloPress,2006,後有 2019 第三版)裡把這件事系統化:7 個訓練強度區間,每個區間對各項生理適應的刺激強度都不同。
六大生理適應
Coggan 整理出的生理適應涵蓋幾個方向:粒線體酶活性、乳酸閾值、肌肉肝醣儲量、血漿容積與心輸出量、肌肉毛細血管密度,以及肌纖維類型轉換。每一項都可以獨立改善,也各有其最有效的訓練強度。
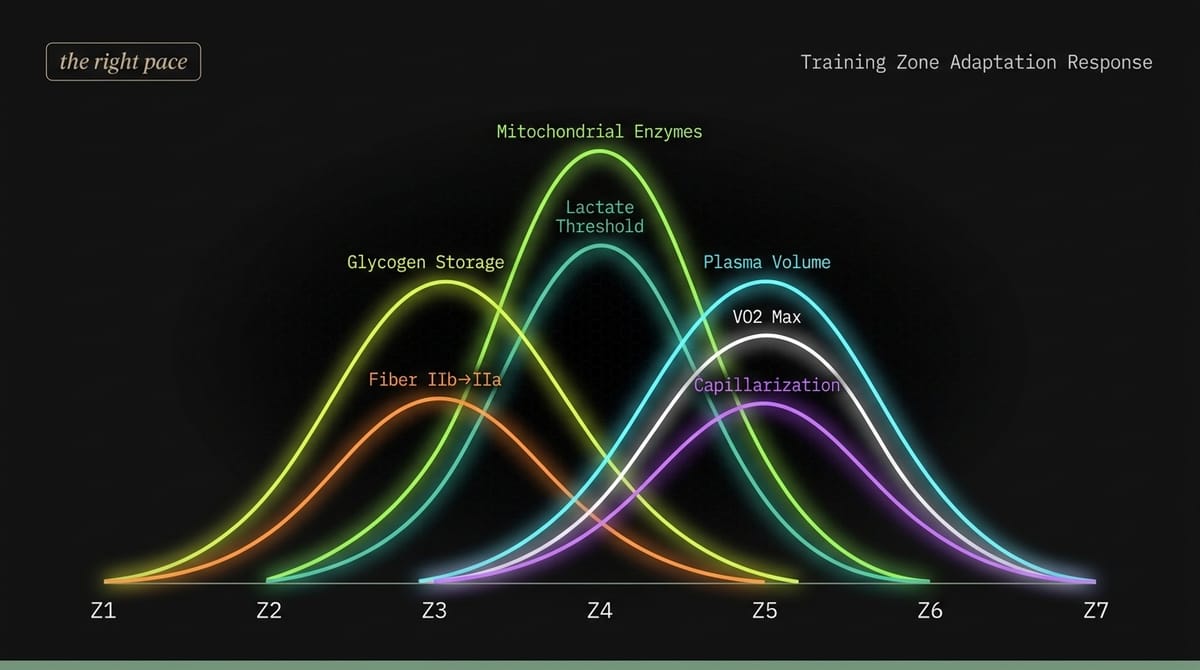
| 生理適應 | Z1 | Z2 | Z3 | Z4 | Z5 | Z6 | Z7 |
|---|---|---|---|---|---|---|---|
| 血漿容積↑ | — | ● | ●● | ●●● | ●●●● | ● | — |
| 肌肉粒線體酶活性↑ | — | ●● | ●●● | ●●●● | ●● | ● | — |
| 乳酸閾值↑ | — | ●● | ●●● | ●●●● | ●● | ● | — |
| 肌肉肝醣儲量↑ | — | ●● | ●●●● | ●●● | ●● | ● | — |
| 慢縮肌纖維肥大 | — | ● | ●● | ●● | ●●● | ● | — |
| 肌肉毛細血管密度↑ | — | ● | ●● | ●● | ●●● | ● | — |
| 快縮肌纖維轉型(IIb → IIa) | — | ●● | ●●● | ●● | ●● | ● | — |
| 心搏量 / 最大心輸出量↑ | — | ● | ●● | ●●● | ●●●● | ● | — |
| VO2max↑ | — | ● | ●● | ●●● | ●●●● | ● | — |
| 肌肉高能磷酸鹽儲量(ATP/PCr)↑ | — | — | — | — | — | ● | ●● |
| 無氧能力↑(乳酸耐受性) | — | — | — | — | ● | ●●● | ● |
| 快縮肌纖維肥大 | — | — | — | — | — | ● | ●● |
| 神經肌肉功率↑ | — | — | — | — | — | ● | ●●● |
●●●● = 峰值刺激 ●●● = 強效 ●● = 中等 ● = 輕微 — = 無顯著效果
看完這張表,有兩件事很清楚。第一,Zone 1(積極恢復)幾乎沒有任何適應效果,它的功能是讓身體從前一次訓練中恢復,不是建設新的能力。第二個可能更出乎意料:Zone 4(閾值)在粒線體酶和乳酸閾值這兩欄的得分最高,不是 Zone 2,也不是 Zone 5。
粒線體:細胞的燃料工廠
粒線體是有氧能量代謝的核心,負責把葡萄糖和脂肪酸氧化成 ATP。訓練之後,粒線體的數量增加、體積增大、氧化酶活性提升——這些變化讓你用同樣的力氣跑得更快,或者用同樣的速度跑得更省力。
這個適應的峰值在 Zone 4(閾值區),Zone 3(Tempo)次之,Zone 5 也有貢獻但效果遞減。機制是:強度夠高時,細胞的能量消耗速度超過補充速度,細胞內的「能量警報」AMPK 被觸發,啟動下游的粒線體增生訊號(PGC-1α)。強度越高,警報越強;但強度太高疲勞積累太快,無法頻繁重複。Zone 4 剛好落在訊號夠強、又能週週重複的甜蜜點上。
那 HIIT 和穩態有氧,哪個對粒線體更有效?單項研究裡,HIIT 組的檸檬酸合酶(粒線體氧化容量的指標酶)增幅更大——高強度每次觸發的訊號確實更強。但 Abrego-Guandique 等人(2025)發現,工作總量相當時,兩者的粒線體適應效果並無顯著差異。觸發粒線體增生的分子路徑相同,HIIT 是每次訊號強但次數少,穩態有氧是訊號溫和但頻率高,積累下來效果接近。
乳酸閾值:把乳酸變成燃料
乳酸閾值速度提升,代表你在更高強度下產生的乳酸,被清除的速度跟上了生產速度。核心機制是乳酸穿梭系統:乳酸在快縮肌裡大量產生,透過 MCT1 轉運蛋白被送到氧化型纖維的粒線體裡燃燒——訓練後 MCT1 密度上升,清除速度加快,累積閾值往更高強度移動。Benítez-Muñoz 等人(2024)確認,MCT1 密度的提升在各種有氧訓練模式中都一致出現。機制的完整說明在前篇〈乳酸不是你的敵人〉裡。
這個適應的峰值在 Zone 4。Zone 4 的強度夠高,能持續給乳酸清除系統施壓,進一步提升 MCT1 密度和酶效率;同時,疲勞積累的速度比 Zone 5 慢,可以更頻繁地重複刺激。這正是挪威法的生理邏輯——Casado 等人 2023 年記錄了 Ingebrigtsen 兄弟每週 3–4 次以血乳酸 2–4.5 mmol/L 為目標的閾值間歇,精算的就是在最大化乳酸閾值適應的同時,維持足夠的訓練頻率。
肌肉肝醣儲量:能量的庫存量
這一項的峰值在 Zone 3(Tempo 區)——在 Coggan 的表格裡,肝醣儲量這一行的最高分落在 Z3,而非 Z4 或 Z5。
機制是:中高強度訓練反覆消耗肌肉肝醣,每次消耗之後的再合成過程,身體會把儲量「超補償」到比訓練前更高的水準。GLUT4 轉運蛋白(負責把葡萄糖送進細胞)的數量也隨訓練提升,肝醣儲存的上限隨之增加。Zone 3 在這個面向的優勢,來自它介於高消耗(夠消耗肝醣)和低疲勞(不需要太長恢復)之間的特性,可以比 Zone 4/5 更頻繁地重複。對馬拉松跑者來說,這直接影響後半段有沒有撞牆。
心肺輸出:管道的容量
VO2max 由兩個部分決定:心臟每分鐘能輸出多少血液,以及肌肉能從這些血液使用多少氧氣。在 Coggan 的表格裡,這兩欄的峰值都在 Zone 5。
Helgerud 等人 2007 年的 RCT 直接驗證了這一點:40 名受試者隨機分成四組,8 週後 Zone 5 間歇組的 VO2max 提升幅度最大,心搏量的增幅與 VO2max 的改善高度吻合;Zone 2 和 Zone 3 組在這兩項指標上均未顯著改善。心臟每次輸出更多血液,才是 VO2max 上升的主要驅動機制,Zone 5 的強度讓這個訊號最強、提升幅度最大。
肌肉毛細血管:氧氣的最後一哩
送出去的血,要能抵達肌肉纖維才算數。毛細血管是那個抵達的管道。訓練之後,肌肉裡的毛細血管密度增加(血管新生,angiogenesis),氧氣從血液擴散到肌肉細胞的距離縮短,利用效率提高。
在 Coggan 的表格裡,這一欄的峰值在 Zone 5。促進血管新生的主要分子信號是 VEGF(血管內皮生長因子)。Wahl 等人 2014 年直接比較高強度訓練和高量低強度訓練的 VEGF 反應——高強度組運動後 VEGF 顯著上升,高量低強度組反而下降。強度越高,代謝缺氧訊號越強,VEGF 反應越大——Zone 5 在這個適應上的效率最高,但並非只有 Zone 5 才能觸發,而是隨強度提升效果遞增。
肌纖維轉換:快縮肌的有氧化
Coggan 的表格把肌纖維適應分成兩個獨立的行。IIb→IIa 轉型的峰值落在 Zone 3(●●●),Zone 2 和 Zone 4/5 效果較低(●●);慢縮肌纖維肥大(slow twitch hypertrophy)則不同,峰值在 Zone 5(●●●),Zone 3/4 中等。兩個看似相近的適應,最強刺激區間差了兩格。Plotkin 等人(2021)確認,長距離訓練後慢縮纖維比例提升,高強度訓練讓更多 IIa 纖維出現——不同強度打造了不同方向的纖維轉換。
無氧能力與乳酸耐受性:Zone 6 的專屬領域
在 Coggan 的表格裡,Zone 6(接近最大強度的短暫爆發,約 30 秒至 2 分鐘)是「無氧能力/乳酸耐受性」這一欄的峰值(●●●)。這個強度讓糖解速率遠超過粒線體的清除能力,乳酸和氫離子快速累積。反覆承受這個刺激,身體的緩衝能力會適應:肌肉和血液中的碳酸氫鹽濃度提升,細胞內 pH 值下降的速度放緩,在高乳酸環境下維持輸出的時間延長。
Zone 6 對一般有氧基礎訓練的貢獻相對低(粒線體、乳酸閾值、VO2max 各欄都只有 ●),但對 800m 至 1500m 這類需要在乳酸大量累積下仍維持配速的項目至關重要。換句話說,Zone 6 不是在打造有氧引擎,而是在訓練引擎過熱時的耐受上限。
神經肌肉功率:Zone 7 的終點站
Zone 7(全力衝刺,通常 10–15 秒以內)在 Coggan 表格裡的峰值落在神經肌肉功率(●●●),同時也是快縮肌纖維肥大(●●)和高能磷酸鹽儲量(ATP-PCr,●●)的重要刺激區間。
機制上,Zone 7 的適應走的是神經路徑,而非代謝路徑:運動單元招募率提升、運動神經元放電頻率加快、肌肉間協調性改善——這些變化讓你能在瞬間動員更多快縮肌纖維。磷酸肌酸(PCr)系統在衝刺頭幾秒提供能量,Zone 7 訓練後 PCr 儲量和再合成速率都會提升。對中長跑選手而言,Zone 7 的貢獻集中在最後衝刺能力,不在比賽主體的有氧輸出。
脂肪燃燒的誤解
「燃脂要練 Zone 2」——這個說法混淆了兩件事:運動當下的能量來源,以及訓練後長期的代謝能力提升。燃脂能力的核心是粒線體的氧化酶活性,特別是 β-HAD(脂肪酸氧化的關鍵酶)。
Talanian 等人(2007)記錄,2 週 HIIT 後,全身脂肪氧化能力提升 36%,粒線體酶 β-HAD 活性上升 32%。機制和粒線體增生走的是同一條路徑:AMPK → PGC-1α → 粒線體酶活性提升 → 脂肪氧化能力增強。HIIT 研究先驅 Martin Gibala 參與的回顧總結:「大多數研究不支持 Zone 2 是提升粒線體容量或脂肪氧化能力的最優強度。」決定燃脂能力的是粒線體容量,不是訓練當下的強度區間。
把上面幾項適應整理在一起,峰值強度的分布很清楚:
- Zone 3(Tempo):肌肉肝醣儲量、IIb→IIa 肌纖維轉型
- Zone 4(閾值):粒線體酶活性、乳酸閾值
- Zone 5(VO2max 區):VO2max、心輸出量、毛細血管密度、慢縮肌纖維肥大
- Zone 6(近最大強度):無氧能力、乳酸耐受性
- Zone 7(全力衝刺):神經肌肉功率、ATP-PCr 儲量、快縮肌纖維肥大
Zone 2 在每個適應欄都有貢獻,但沒有任何一項的峰值落在 Zone 2。它的優勢是疲勞低、可以累積大量訓練量——量夠高的時候,溫和的刺激也能積累出可觀的效果。Zone 2 是累積訓練量的工具,不是觸發峰值適應的工具。
訓練法:殊途同歸
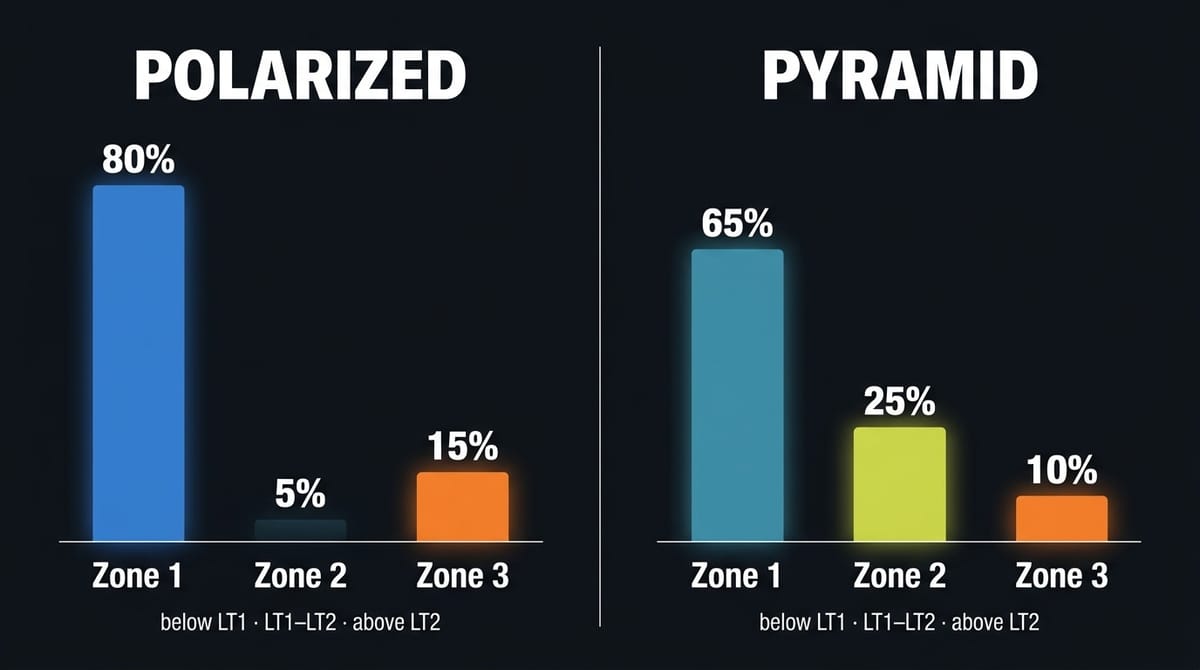
以三區模型來看:極化訓練(80/20 框架)把約 80% 的時間壓在第一區(LT1 以下的低強度),約 20% 投入第三區(LT2 以上的高強度),中間的第二區(LT1 到 LT2 之間的節奏到閾值強度)佔比極低。金字塔訓練讓強度分布呈遞減結構:大量第一區累積有氧基礎,一定比例的第二區閾值刺激,少量第三區觸發上限適應。
哪個更好?Oliveira 等人(2024)比較了極化與其他強度分配模型(17 項 RCT,n=437):時間試驗成績、力竭時間、LT2 速度,全無顯著差異。極化訓練在 VO2max 上略有優勢,但這個效果只在短期介入(< 12 週)和高水準運動員身上出現,長期介入後差距幾乎消失。
不同訓練法優先打造不同的生理零件:極化偏重 VO2max 和心輸出量,閾值訓練偏重粒線體酶和乳酸閾值,高量低強度偏重脂肪氧化基礎。最終成績取決於哪些生理機轉是你的瓶頸,不是走哪條路徑。
跑多少,跑多快,不是二選一
Fokkema 等人 2020 年追蹤近千名跑者備賽半馬與全馬的訓練資料,結果很清楚:週跑量和訓練配速都獨立預測完賽時間,且各自的貢獻互不重疊。低跑量加高配速和高跑量加較低配速,激活的生理適應比例不同,但有氧負荷夠高時,競技水準可以相近。
這也解釋了為什麼頂尖跑者進行不同的訓練方式,卻都能在奧運決賽跑道上相遇。
每個跑者走的路不一樣,但讓你進步的生理機制是相同的。
路不決定你跑多快,你在這條路上走了多少,才決定。
參考文獻
- 1. Allen, H., & Coggan, A. R. (2019). Training and racing with a power meter (3rd ed.). VeloPress. (Original work published 2006)
- 2. Benítez-Muñoz, J. A., Rubio-Arias, J. Á., Valls-Lacalle, L., González-Ponce, Á., de Vera-Floristán, C., López-Ayala, J. M., & Jiménez, S. L. (2024). Exercise influence on monocarboxylate transporter 1 (MCT1) and 4 (MCT4) in the skeletal muscle: A systematic review. Acta Physiologica, 240(1), e14083. ↗
- 3. Casado, A., Foster, C., Bakken, M., & Tjelta, L. I. (2023). Does lactate-guided threshold interval training within a high-volume low-intensity approach represent the "next step" in the evolution of distance running training? International Journal of Environmental Research and Public Health, 20(5), 3782. ↗
- 4. Helgerud, J., Høydal, K., Wang, E., Karlsen, T., Berg, P., Bjerkaas, M., Simonsen, T., Helgesen, C., Hjorth, N., Bach, R., & Hoff, J. (2007). Aerobic high-intensity intervals improve VO2max more than moderate training. Medicine & Science in Sports & Exercise, 39(4), 665–671. ↗
- 5. Wahl, P., Jansen, F., Achtzehn, S., Schmitz, T., Bloch, W., Mester, J., & Werner, N. (2014). Effects of high intensity training and high volume training on endothelial microparticles and angiogenic growth factors. PLOS ONE, 9(5), e96024. ↗
- 6. Talanian, J. L., Galloway, S. D. R., Heigenhauser, G. J. F., Bonen, A., & Spriet, L. L. (2007). Two weeks of high-intensity aerobic interval training increases the capacity for fat oxidation during exercise in women. Journal of Applied Physiology, 102(4), 1439–1447. ↗
- 7. Oliveira, P. S., Boppre, G., & Fonseca, H. (2024). Comparison of polarized versus other types of endurance training intensity distribution on athletes' endurance performance: A systematic review with meta-analysis. Sports Medicine, 54, 2817–2835. ↗
- 8. Fokkema, T., van Damme, A. A. D. N., Fornerod, M. W. J., de Vos, R.-J., Bierma-Zeinstra, S. M. A., & van Middelkoop, M. (2020). Training for a (half-)marathon: Training volume and longest endurance run related to performance and running injuries. Scandinavian Journal of Medicine & Science in Sports, 30(9), 1692–1704. ↗
- 9. Storoschuk, K. L., Moran-MacDonald, A., Gibala, M. J., & Gurd, B. J. (2025). Much ado about zone 2: A narrative review assessing the efficacy of zone 2 training for improving mitochondrial capacity and cardiorespiratory fitness in the general population. Sports Medicine, advance online. ↗
如果這篇文章對你有幫助,歡迎 請我喝杯咖啡 ☕ 繼續創作。