Running 100 meters is “anaerobic,” running a marathon is “aerobic” — you’ve heard this before. It’s convenient, intuitive, and almost completely wrong.
Inside real muscle, three energy systems never take turns. They all run simultaneously, with proportions shifting as intensity changes.
What Is ATP
Muscles need energy to contract, but they can’t burn food directly. Food must first be converted into ATP (adenosine triphosphate) — think of it as the body’s universal energy currency. Muscles only accept this format.
The problem is that muscles store very little ATP — all-out effort exhausts the supply in seconds. The job of all three energy systems is to continuously replenish the ATP that gets used. The faster and more abundantly they replenish it, the higher the intensity you can sustain for longer.
Three Systems, Each with Its Role
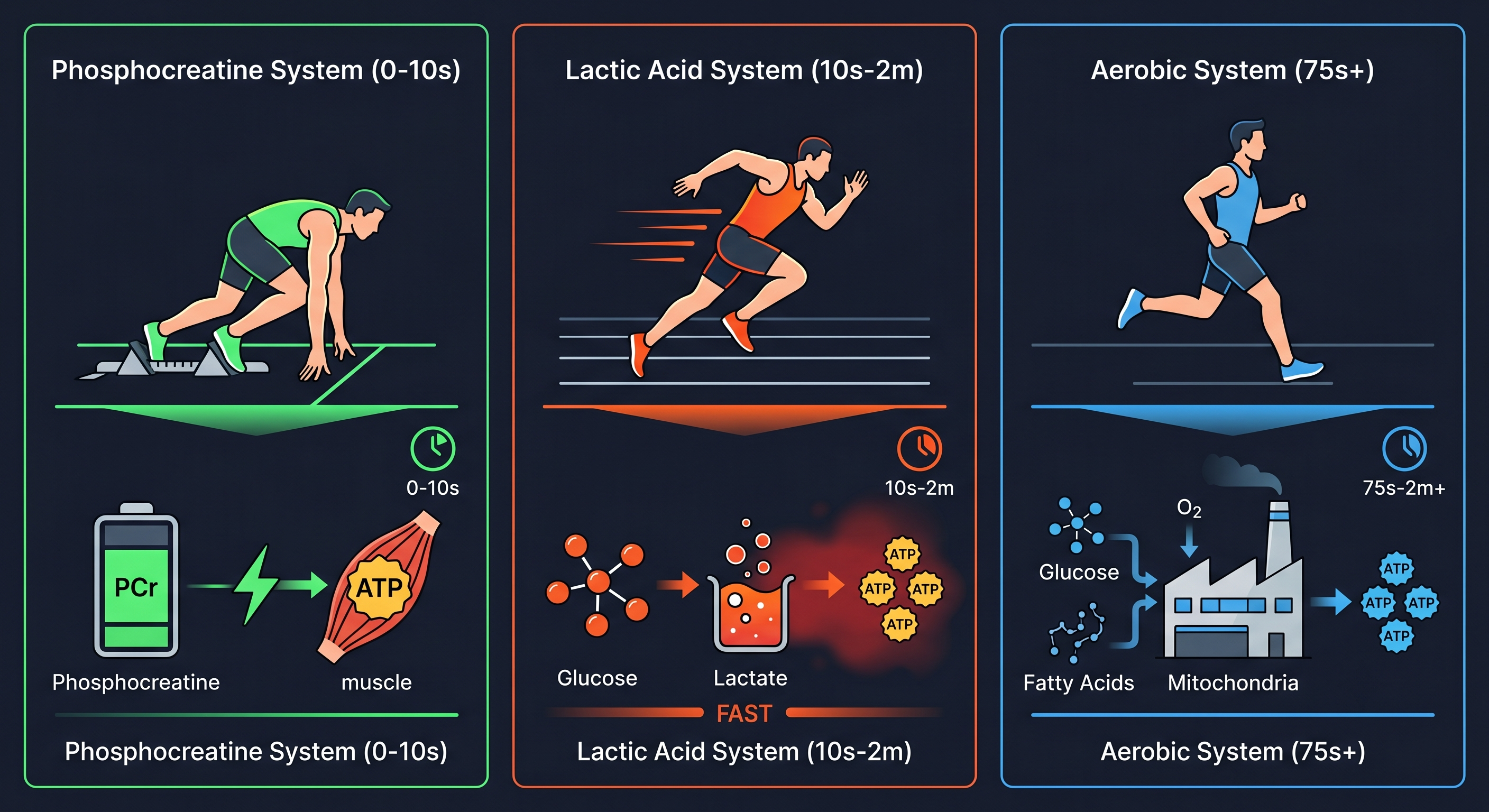
The fastest system. Alongside ATP, muscles store phosphocreatine (PCr) — think of it as a backup battery. The moment ATP runs low, PCr plugs in and recharges it almost instantly, with no oxygen required.
The tradeoff: very limited reserves. Within 10 seconds of high-intensity effort, more than half is depleted; within a few minutes, nearly all of it is gone. The 100-meter sprint relies primarily on this system, but it can’t sustain prolonged effort.
Lactate System: 10 seconds to about 2 minutes
As PCr reserves fall, the body shifts to glycolysis — breaking down glucose into lactate at a rate roughly 100 times faster than the aerobic system, quickly bridging the energy gap when intensity spikes.
The tradeoff: low efficiency. Each glucose molecule yields only 2 ATP; the aerobic pathway produces 32. Fast, but costly. The 400- to 800-meter races are where this system does most of the work.
Aerobic System: Active from the Start, Dominant after 75 Seconds
The slowest to ramp up, but with near-unlimited capacity. Glucose and fat are fully oxidized in the mitochondria — one glucose yields about 32 ATP, fat yields even more, and it runs continuously as long as there’s oxygen and fuel. Marathon pace is almost entirely sustained by this system.
Its weakness is slow activation. When high-intensity demand spikes suddenly, the aerobic system can’t keep up — that’s why the first two systems step in to bridge the gap. What limits the aerobic system isn’t fuel; it’s the rate at which oxygen can be delivered.
Three Systems Running at the Same Time
None of these systems ever “turn off.”
Research has measured the energy contribution of each system at different exercise intensities:
| Duration | Phosphocreatine | Lactate System | Aerobic System |
|---|---|---|---|
| 10 sec (full sprint) | 53% | 44% | 3% |
| 30 sec | 23% | 49% | 28% |
| 75+ sec | Declining | Peaking then declining | Above 50%, still rising |
During a full-effort 10-second sprint, the aerobic system is still contributing 3% — it never fully stops. By 30 seconds, the aerobic share has already jumped to 28%.
This means there is no such thing as “purely anaerobic running” — only “running with a higher anaerobic proportion.” Conversely, even at marathon pace, the phosphocreatine system never fully shuts down; it contributes to every push-off step.
Intensity changes the ratio — not the switch.
Fast-Twitch and Slow-Twitch Muscle
Before talking about lactate, there’s a foundational concept to establish: not all muscle fibers are the same. Because lactate circulates between fiber types, the next section won’t make sense without this.
Slow-twitch fibers (Type I): aerobically powerful, packed with mitochondria, fatigue-resistant, built for sustained output. The workhorse of long-distance running.
Fast-twitch fibers (Type II): high force, high power, but weaker oxidative capacity, relatively quick to fatigue. Sprinting, jumping, and explosive movements rely on these.
This difference directly shapes how the lactate system behaves.
Lactate Is Fuel
Many people believe running fatigue is caused by lactate accumulation. That picture needs updating.
The lactate system in fast-twitch fibers continuously produces lactate — not only at high intensity, but even during easy jogging, fast-twitch fibers are running glycolysis and generating lactate. What happens then is that this lactate travels through the blood to neighboring slow-twitch fibers, where it’s taken up into the mitochondria and oxidized as fuel for the aerobic system. At low intensity, the rate of lactate production roughly matches the rate of utilization, so blood concentration doesn’t build up.
The heart also preferentially takes up lactate as fuel. During high-intensity exercise, lactate can supply up to 60% of cardiac muscle’s energy needs.
Lactate is not a waste product — it’s a circulating fuel. Rising blood lactate is a signal that the body is running efficiently, with fast-twitch fiber production outpacing slow-twitch consumption. The mechanism that makes your legs give out in the late stages of a race is a different story, and its relationship to lactate itself is far less direct than traditionally assumed.
(A full research breakdown on this topic is in Lactate Is Not Your Enemy.)
The Design of Fatigue
When muscles reach exhaustion during all-out effort, ATP doesn’t actually hit zero.
At rest, ATP concentration is around 8 mmol/kg. At the point of complete collapse, the minimum is roughly 5 mmol/kg. Energy demand may spike 1000-fold, but ATP drops by only a few units. This isn’t accidental — it’s a protective mechanism. If ATP truly ran out, actin and myosin in the muscle fibers would lock up, unable to relax, and the muscle would enter rigor.
The point at which your legs stop working is the body applying the brakes before an ATP crisis can occur. It stops you — before it breaks you.
From this perspective, fatigue is a precision protection system, not a sign of failure.
Training Is Mostly About Upgrading the Aerobic Engine
Of the three systems, training produces the most visible adaptations in the aerobic system.
Mitochondrial density, efficiency, and the electron transport chain capacity within each mitochondrion all change with endurance training. Six months of endurance training can increase mitochondrial maximum ATP production rate (ATPmax) by 32%, with a corresponding 17% increase in leg power output. This improvement is roughly evenly distributed across three directions: more mitochondria, higher efficiency per mitochondrion, and greater electron transport chain capacity.
From rest to all-out effort, the aerobic system’s ATP output can span a 50-fold range. The larger that range, the more energy is available across every intensity level.
Once the aerobic foundation improves, it’s not just easy running that gets faster — high-intensity capacity also advances. This is because the lactate system is under less pressure, and the phosphocreatine system recovers faster. The three systems are not independent: build the aerobic base solidly, and both of the other systems perform better.
Once You Understand This, Training Logic Follows Naturally
Why Is Zone 2 the Foundation of Endurance?
Zone 2 is the intensity range where the aerobic system is highly engaged, but the rate of lactate accumulation hasn’t yet exceeded the rate of utilization — roughly the pace where you can still form complete sentences, but have started to feel the effort. At this intensity, slow-twitch fibers work continuously, and mitochondria receive sustained, stable stimulus, gradually increasing in both number and efficiency.
Many runners find Zone 2 “too easy — nothing’s happening,” and push the intensity up. But once the lactate system is driving, the direction of adaptation shifts. Zone 2 is about upgrading the mitochondrial factory, and that adaptation requires accumulated time — not acute suffering.
Rest Interval Length in Interval Training Determines Which System You’re Training
Full recovery of phosphocreatine requires 5 to 15 minutes. Short sprint intervals (6 to 10 seconds all-out) paired with sufficient rest allow the phosphocreatine system to start each rep with full reserves — the training target is that system’s explosive power.
If the goal is to stimulate the lactate system, shortening rest is deliberate — keeping blood lactate elevated and forcing the lactate system to operate under sustained metabolic pressure. That’s the logic behind 400- to 800-meter pace intervals.
Rest intervals aren’t “longer is better” — they’re part of the training target. You design the combination that puts maximum stress on the system you want to strengthen.
Why Does Training Only at Race Pace Lead to a Plateau?
Without raising the ceiling of the aerobic engine, you hit a speed wall quickly. You can push through with willpower, but the cost is a lactate system that’s overloaded every session — slower recovery, and cumulative fatigue that outpaces adaptation. With a weak aerobic base, speed training can only deliver gains within a narrow window.
This is also why many runners who have trained seriously for years start improving only after they add a large volume of easy running — it’s not that slow running makes you faster. It’s that upgrading the aerobic engine raises the returns on training across the board.
Energy is not three separate lines. It’s a flame that always burns simultaneously, with the proportions constantly in motion.
References
- 1. Spriet, L. L. (2012). Interaction among skeletal muscle metabolic energy systems during intense exercise. Journal of Nutrition and Metabolism, 2012, 905612. ↗
- 2. Adeva-Andany, M. M., et al. (2014). Biochemistry, anaerobic glycolysis. StatPearls. ↗
- 3. Layec, G., et al. (2016). Mitochondria to motion: optimizing oxidative phosphorylation to improve exercise performance. Journal of Physiology, 594(18), 5231–5241. ↗
- 4. Harms, M., & Johansson, A. (2022). A century of exercise physiology: key concepts on coupling respiratory oxygen flow to muscle energy demand during exercise. European Journal of Applied Physiology. ↗
If this article was helpful, feel free to buy me a coffee ☕ and keep me writing.